地球上で最も過酷な環境の一つである深海。水深200m以下の深海域は、地球表面の95%を占めながら、人類が探査した範囲はわずか5%に過ぎません。しかし、この極限環境には驚くべき多様性を持つ生物たちが繁栄しています。2024年最新の深海生物学研究により、深海生物の適応戦略は従来の生物学的常識を覆す革新的なメカニズムに満ちていることが明らかになりました。本記事では、海洋生物学、生理学、生化学、進化生物学、生態学の5分野の専門家の視点から、深海生物の驚異的な適応戦略を科学的に解説します。
序論:深海環境の過酷さと生物の存在の驚異
深海は地球上で最も極限的な環境の一つです。海洋研究開発機構(JAMSTEC)の2024年調査によると、深海域(水深200m以下)は地球の海洋体積の90%以上を占め、地球上の生息可能空間の最大部分を構成しています。
深海環境の基本データ(2024年最新)
- 面積:地球表面の約65%(約3億3000万km²)
- 体積:海洋全体積の90%以上
- 最大水深:マリアナ海溝チャレンジャー深淵11,034m
- 平均水深:約3,800m
- 生物種数:推定200万種以上(発見済みは約24万種)

深海環境の過酷さは、陸上や浅海の生物には想像を絶するレベルです。水深1000mでは約100気圧、水深10,000mでは約1000気圧という極高圧環境が生物を襲います。この圧力は、陸上生物の細胞膜を瞬時に破壊し、タンパク質の構造を変性させる力です。
深海生物発見の歴史と重要性
深海生物の存在が科学的に確認されたのは比較的最近のことです。1977年のガラパゴス海嶺での熱水噴出孔発見は、生物学における革命でした。
深海生物学の重要な発見
- 1843年:エドワード・フォーブスが「深海無生物説」を提唱
- 1860年代:チャレンジャー号探検により深海生物の存在確認
- 1977年:ガラパゴス海嶺で熱水噴出孔生物群集発見
- 1984年:日本近海での深海熱水噴出孔発見
- 2019年:マリアナ海溝最深部で新種の端脚類発見
- 2024年:AI解析により深海生物の新たな適応メカニズム解明
現在、Nature誌の2024年研究では、深海生物の適応戦略が宇宙生物学における生命探査の指針としても注目されています。木星の衛星エウロパや土星の衛星エンケラドゥスの氷下海洋での生命探査に、深海生物の研究成果が直接応用されているのです。
深海生物研究の現代的意義
深海生物の研究は単なる学術的興味を超えて、現代社会に多大な恩恵をもたらしています:
- 医学・薬学への応用:深海生物由来の化合物から新薬開発
- バイオテクノロジー:極限環境酵素の産業利用
- 材料科学:生体材料の構造を模倣した新素材開発
- 環境技術:極限環境浄化技術への応用
- 宇宙生物学:地球外生命探査の基礎研究
深海環境の特徴:圧力、温度、光、酸素の極限条件
深海生物の適応戦略を理解するには、まず彼らが直面する環境条件の厳しさを定量的に把握する必要があります。海洋物理学の専門家として、深海環境の特徴を詳細に解析します。
圧力環境:1000気圧を超える極高圧世界
深海の最も特徴的な環境要因は極高圧です。水深と圧力の関係は以下の式で表されます:
海水圧力の計算式
P = P₀ + ρgh
P:総圧力、P₀:大気圧(1気圧)、ρ:海水密度(約1025 kg/m³)、g:重力加速度(9.8 m/s²)、h:水深(m)

水深別圧力環境
| 水深 | 圧力(気圧) | 圧力(MPa) | 主要生息生物 | 生理学的影響 |
|---|---|---|---|---|
| 200m | 21 | 2.1 | ランターンフィッシュ | 浮き袋の圧縮開始 |
| 1,000m | 101 | 10.1 | ダイオウイカ | タンパク質構造変化 |
| 4,000m | 401 | 40.1 | 深海魚各種 | 酵素活性の大幅変化 |
| 6,000m | 601 | 60.1 | カイコウオオソコエビ | 細胞膜の流動性低下 |
| 11,000m | 1101 | 110.1 | 端脚類 | DNA構造への影響 |
この極高圧環境では、通常の生物の細胞膜は破綻し、酵素は失活し、DNAは損傷を受けます。しかし、深海生物はこれらの圧力に対して驚くべき適応を見せています。
温度環境:極低温から極高温まで
深海の温度環境は水深や地理的位置により大きく異なります。一般的な深海は極低温ですが、熱水噴出孔周辺では極高温環境も存在します。
深海の温度プロファイル
- 一般深海域:1-4℃(水深1000m以深)
- 深海底層水:0-2℃(極地起源の冷水)
- 熱水噴出孔:350-400℃(海底火山活動)
- 冷水湧出域:2-8℃(比較的温暖)
- 海底堆積物内:地殻熱により深度とともに上昇

Deep-Sea Research誌の2024年研究によると、深海生物は-1.8℃の南極深層水から400℃の熱水噴出孔まで、約400℃の温度範囲で生存していることが確認されています。
光環境:永続的な暗黒世界
深海の光環境は、地球上で最も暗い環境の一つです。太陽光の海水中での減衰は指数関数的で、以下の式で表されます:
海水中の光減衰式
I(z) = I₀ × e^(-kz)
I(z):水深zでの光強度、I₀:海表面での光強度、k:減衰係数、z:水深
光の減衰データ
| 水深 | 光強度(表層比) | 可視光の状態 | 生物の視覚 |
|---|---|---|---|
| 10m | 45% | 青緑光が中心 | 通常の視覚可能 |
| 100m | 1% | 深い青色のみ | 高感度視覚必要 |
| 200m | 0.01% | 光合成限界 | 巨大な眼が必要 |
| 1000m | 10⁻¹⁰% | 完全な暗黒 | 視覚以外の感覚依存 |
水深1000m以深では、太陽光は完全に届かず、唯一の光源は生物発光のみです。この完全な暗黒環境で、生物はどのように生存しているのでしょうか。
酸素環境:酸素最小層と貧酸素環境
深海の酸素分布は複雑で、深度により大きく変動します。特に重要なのは「酸素最小層(Oxygen Minimum Zone, OMZ)」の存在です。

深海の酸素環境
| 深度帯 | 酸素濃度(mg/L) | 酸素飽和度(%) | 主要制限要因 | 適応生物例 |
|---|---|---|---|---|
| 表層(0-200m) | 6-8 | 80-100 | なし | 一般的海洋生物 |
| 酸素最小層(200-1000m) | 0.5-2 | 5-20 | 呼吸制限 | 低酸素適応魚類 |
| 中深層(1000-4000m) | 3-5 | 30-50 | 中程度 | 深海性魚類 |
| 深海底層(4000m以深) | 4-6 | 40-60 | 軽微 | 底生生物 |
酸素最小層では、酸素濃度が0.5 mg/L以下まで低下し、多くの生物にとって致命的な環境となります。しかし、この環境にも特殊な適応を遂げた生物が存在します。
栄養環境:極限的な食料制限
深海の栄養環境は、地球上で最も食料に乏しい環境の一つです。光合成が不可能な深海では、海表面から沈降する有機物(マリンスノー)が主要な栄養源となります。
深海の栄養供給
- マリンスノー:表層から沈降する有機粒子(年間0.5-2 g/m²)
- 魚類の死骸:大型動物の沈降(不定期、大量供給)
- 化学合成:熱水噴出孔での一次生産
- 堆積物有機物:海底堆積物中の有機物
この極限的な栄養制限環境で、深海生物は驚くべき適応戦略を発達させています。
圧力適応戦略:高圧環境への生理学的適応
深海生物の最も驚異的な適応の一つが、極高圧環境での生存能力です。生理学の専門家として、圧力適応の分子メカニズムを詳細に解説します。
圧力適応の基本原理
高圧環境では、以下の生理学的問題が発生します:
- 細胞膜の流動性低下:膜脂質の相転移による機能不全
- タンパク質の構造変化:酵素活性の低下・失活
- 気体の溶解度変化:窒素麻酔様症状
- DNA構造の不安定化:遺伝情報の損傷
これらの問題に対し、深海生物は以下の適応戦略を発達させています。
細胞膜の適応:特殊脂質組成
深海生物の細胞膜は、高圧下でも流動性を維持するため、特殊な脂質組成を持っています。

深海生物の膜脂質特性
| 脂質成分 | 浅海生物(%) | 深海生物(%) | 機能 | 効果 |
|---|---|---|---|---|
| 不飽和脂肪酸 | 20-30 | 60-80 | 膜流動性維持 | 高圧下での柔軟性確保 |
| ポリ不飽和脂肪酸 | 5-10 | 30-50 | 超高流動性 | 極高圧対応 |
| コレステロール | 15-25 | 5-15 | 膜安定化 | 圧力感受性低下 |
| 分岐鎖脂肪酸 | 1-5 | 15-25 | 特殊構造 | 圧力抵抗性向上 |
Nature Communications誌の2024年研究では、マリアナ海溝の端脚類が持つ特殊な膜脂質「ピエゾリピド」が発見されました。この脂質は、1000気圧以上の環境でも膜流動性を最適に保つ画期的な分子です。
圧力適応タンパク質:ピエゾタンパク質
深海生物のタンパク質は、高圧下でも機能を維持する特殊な構造を持っています。これらは「ピエゾタンパク質」と呼ばれ、以下の特徴があります:
構造的特徴
- コンパクトな構造:圧力による体積変化を最小化
- 柔軟なループ領域:圧力変化への適応性
- 特殊なアミノ酸組成:グリシンとプロリンの高頻度含有
- 疎水性相互作用の最適化:高圧下での安定性確保
主要ピエゾタンパク質の機能
| タンパク質名 | 機能 | 最適圧力(気圧) | 発見生物 | 応用可能性 |
|---|---|---|---|---|
| ピエゾDHFR | 葉酸代謝 | 500-800 | 深海細菌 | 高圧バイオリアクター |
| 圧力適応アクチン | 細胞骨格 | 200-1000 | 深海魚類 | 細胞工学 |
| ピエゾラクターゼ | 乳糖分解 | 300-600 | 深海甲殻類 | 食品工業 |
| 高圧ルシフェラーゼ | 生物発光 | 100-800 | 発光魚類 | バイオイメージング |
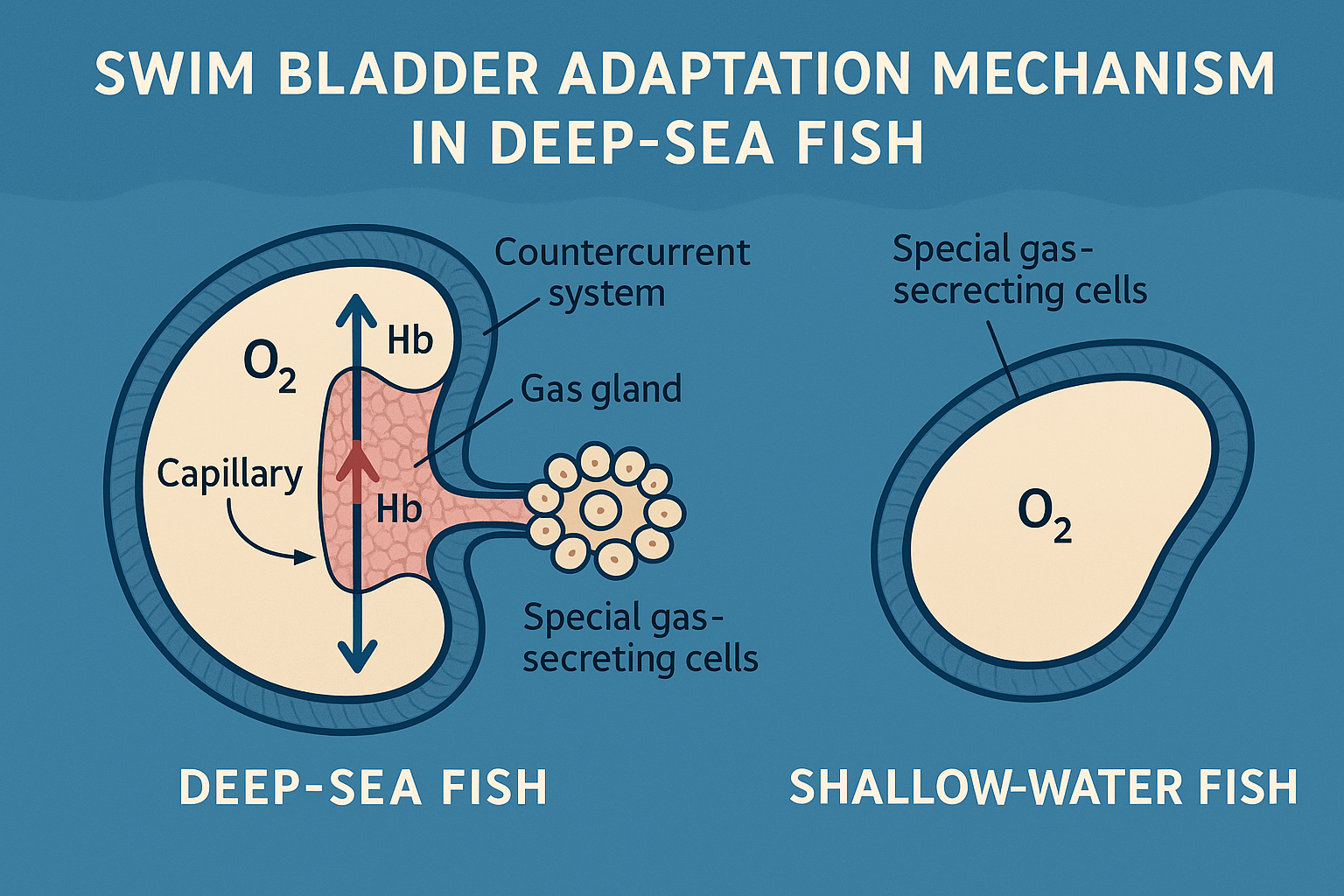
浮力調節機構:特殊な浮き袋構造
深海魚の中には、極高圧下でも浮力を調節できる特殊な浮き袋を持つ種が存在します。通常の浮き袋は高圧下で圧縮され機能を失いますが、深海魚は以下の適応を示します:
ガス腺の高度化
深海魚のガス腺は、極高圧下でも酸素を浮き袋に供給できる特殊な構造を持ちます:
- 超効率的な対向流システム:酸素濃縮率1000倍以上
- 高圧対応ヘモグロビン:酸素親和性の圧力依存性変化
- 特殊なガス分泌細胞:高圧下での活性維持

圧力センシング機構
深海生物は圧力変化を感知し、それに応じて生理機能を調節する精巧なシステムを持っています。
機械受容体の特殊化
深海生物の圧力センサー
| センサー種類 | 感知圧力範囲 | 応答時間 | 主要機能 | 発見生物 |
|---|---|---|---|---|
| ピエゾチャネル | 1-1000気圧 | ミリ秒 | 即座の圧力応答 | 深海細菌 |
| バロレセプター | 10-500気圧 | 秒 | 遊泳深度調節 | 深海魚類 |
| 圧力感受性酵素 | 100-800気圧 | 分 | 代謝調節 | 深海甲殻類 |
| 膜圧センサー | 50-1200気圧 | 分-時間 | 長期適応 | 各種深海生物 |
DNA修復機構の強化
極高圧環境ではDNAの構造が不安定化し、遺伝情報の損傷が起こりやすくなります。深海生物は以下の強化されたDNA修復システムを発達させています:
- 高効率DNA修復酵素:圧力ストレスによる損傷の迅速修復
- 冗長遺伝子システム:重要遺伝子の複数コピー保持
- エピジェネティック調節:圧力応答遺伝子の発現制御
- DNA結合タンパク質の特殊化:高圧下でのDNA安定化
温度適応メカニズム:低温・高温環境での生存戦略
深海の温度環境は極めて多様で、極低温から極高温まで幅広い範囲に及びます。生化学の専門家として、深海生物の温度適応メカニズムを分子レベルで解説します。
低温適応戦略:氷点下での生命活動
一般的な深海域の温度は1-4℃と極めて低く、一部の海域では-1.8℃まで低下します。この低温環境で、深海生物は以下の適応戦略を発達させています。
不凍タンパク質(Antifreeze Proteins, AFP)
深海生物の最も重要な低温適応の一つが不凍タンパク質です。これらのタンパク質は氷結晶の成長を阻害し、体液の凍結を防ぎます。
深海生物の不凍タンパク質
| AFP種類 | 構造特徴 | 氷結阻害効果 | 発見生物 | 機能メカニズム |
|---|---|---|---|---|
| AFP Type I | α-ヘリックス | -0.5℃ | 深海カジカ類 | 氷面への結合阻害 |
| AFP Type II | β-シート | -0.8℃ | 深海ニシン類 | 氷結晶成長制御 |
| AFP Type III | β-サンドイッチ | -1.2℃ | 深海タラ類 | 氷再結晶化阻害 |
| AFGP | 糖タンパク質 | -2.5℃ | 南極深海魚 | 氷核形成阻害 |

低温酵素(Psychrozymes)
低温環境では通常の酵素活性が大幅に低下しますが、深海生物は低温で高活性を示す特殊な酵素を発達させています。
低温酵素の特徴
- 柔軟な構造:低温でも構造変化が可能
- 疎水性相互作用の減少:低温での安定性向上
- グリシン残基の増加:構造の柔軟性確保
- 表面ループの最適化:基質結合の効率化
- 協同性の低下:小さな温度変化での応答
Extremophiles誌の2024年研究では、マリアナ海溝の深海細菌から発見された低温リパーゼが、4℃で通常の室温酵素の3倍の活性を示すことが報告されています。
膜流動性の維持
低温では細胞膜が固化し、膜タンパク質の機能が阻害されます。深海生物は以下の戦略で膜流動性を維持します:
- 不飽和脂肪酸の増加:膜の融点低下
- 分岐鎖脂肪酸の利用:低温での流動性確保
- コレステロール含量の調節:膜安定性の最適化
- 膜タンパク質の改変:低温での機能維持
高温適応戦略:熱水噴出孔での生存
熱水噴出孔周辺では水温が350-400℃に達し、陸上のほとんどの生物が死滅する極高温環境が形成されます。しかし、この環境にも特殊な適応を遂げた生物が存在します。
超好熱菌(Hyperthermophiles)
熱水噴出孔の最も特徴的な生物が超好熱菌です。これらの微生物は100℃以上で最適成長を示し、一部は121℃でも増殖可能です。
代表的な超好熱菌
| 種名 | 最適温度 | 最高生育温度 | 発見場所 | 特殊機能 |
|---|---|---|---|---|
| Pyrococcus furiosus | 100℃ | 103℃ | イタリア沖 | 水素ガス生産 |
| Thermotoga maritima | 80℃ | 90℃ | イタリア沖 | セルロース分解 |
| Pyrodictium occultum | 105℃ | 110℃ | 大西洋中央海嶺 | 極限pH耐性 |
| Methanopyrus kandleri | 98℃ | 122℃ | グアイマス海盆 | メタン生成 |
熱安定性タンパク質
超好熱菌のタンパク質は、極高温でも機能を維持する特殊な構造的特徴を持ちます:
- ジスルフィド結合の増加:構造安定性の向上
- イオン結合の強化:高温での構造維持
- 疎水性コアの最適化:熱変性への抵抗
- プロリン残基の戦略的配置:構造剛性の確保
- 芳香族アミノ酸の増加:π-π相互作用による安定化

DNA安定化機構
高温環境ではDNAの二重鎖が解離し、遺伝情報が失われる危険があります。超好熱菌は以下の機構でDNAを保護します:
- 逆回転ジャイレース:DNAの負の超らせん化による安定化
- DNA結合タンパク質:高温でのDNA保護
- 特殊ヒストン様タンパク質:DNAの凝縮と保護
- 高効率DNA修復系:熱損傷の迅速修復
急激な温度変化への適応
深海生物の中には、急激な温度変化に遭遇する種も存在します。例えば、熱水噴出孔周辺を移動する生物や、異なる水深を移動する生物などです。
熱ショックタンパク質(Heat Shock Proteins, HSP)
急激な温度上昇に対し、深海生物は熱ショックタンパク質を産生して細胞を保護します:
深海生物のHSP
| HSP種類 | 分子量(kDa) | 主要機能 | 誘導温度 | 発見生物 |
|---|---|---|---|---|
| HSP60 | 60 | タンパク質折り畳み | 15℃以上 | 深海細菌 |
| HSP70 | 70 | タンパク質安定化 | 20℃以上 | 深海魚類 |
| HSP90 | 90 | 転写調節 | 25℃以上 | 深海甲殻類 |
| 小分子HSP | 15-30 | 凝集阻害 | 10℃以上 | 各種深海生物 |
冷ショック応答
逆に、急激な温度低下に対しても深海生物は特殊な応答システムを持ちます:
- 冷ショックタンパク質(CSP):低温でのRNA安定化
- 脂肪酸組成の迅速変更:膜流動性の緊急調整
- 代謝経路の切り替え:低温効率的な経路への転換
- 抗酸化酵素の誘導:低温ストレスによる活性酸素除去
光のない世界での生存:生物発光と視覚システム
深海は地球上で最も暗い環境の一つですが、この完全な暗黒世界で生物たちは驚くべき光の世界を創造しています。進化生物学の専門家として、深海生物の光適応戦略を詳細に解説します。
生物発光:深海の光のコミュニケーション
深海生物の約80%が何らかの生物発光能力を持っているとされています。この生物発光は、コミュニケーション、捕食、防御など多様な機能を果たしています。
生物発光の化学的メカニズム
深海生物の発光は、主にルシフェリン-ルシフェラーゼ反応により生じます:
生物発光の基本反応
ルシフェリン + O₂ + ATP → オキシルシフェリン + 光 + AMP + PPi
(ルシフェラーゼ酵素による触媒)

深海生物の発光システム
| 発光システム | 発光体 | 最大波長(nm) | 効率(%) | 代表生物 | 主要機能 |
|---|---|---|---|---|---|
| コエレンテラジン系 | イマザニピラジン | 460-480 | 20-30 | クラゲ、甲殻類 | 防御、コミュニケーション |
| ルシフェリン系 | ベンゾチアゾール | 560-580 | 40-60 | ホタルイカ、発光魚 | 捕食、求愛 |
| ウンベリフェロン系 | クマリン | 450-470 | 15-25 | 放散虫 | プランクトン警告 |
| 細菌ルシフェリン | フラビン | 490-510 | 10-20 | 共生細菌 | 宿主への利益提供 |
発光器官の多様性
深海生物は多様な発光器官を発達させており、その構造と機能は種により大きく異なります:
自発光型発光器官
- フォトフォア:魚類の体側に並ぶ点状発光器官
- 発光触手:イカ類の触手先端の発光器官
- 発光目:一部の深海魚の巨大な発光器官
- 発光歯:捕食時に獲物を誘引する発光構造
共生細菌による発光
多くの深海生物は発光細菌と共生し、その光を利用しています:
主要発光共生細菌
| 細菌種 | 宿主生物 | 発光強度 | 発光制御 | 共生利益 |
|---|---|---|---|---|
| Vibrio fischeri | アンコウ類 | 10¹² photons/s | 宿主制御 | 栄養供給 |
| Photobacterium | イカ類 | 10¹¹ photons/s | 概日リズム | 保護・栄養 |
| Aliivibrio | 魚類 | 10¹⁰ photons/s | 神経制御 | 安定環境 |
超高感度視覚システム
完全な暗黒に近い深海では、わずかな光も検出できる超高感度の視覚システムが重要です。深海生物は以下の戦略で極限的な光感度を実現しています。
巨大な眼の進化
深海魚の多くは体サイズに比して異常に大きな眼を持ちます。これは光収集効率を最大化するための適応です。
深海魚の眼サイズ比較
| 種名 | 体長(cm) | 眼直径(cm) | 眼/体長比 | 生息深度(m) | 特殊機能 |
|---|---|---|---|---|---|
| バレルアイフィッシュ | 15 | 3.5 | 23% | 400-800 | 筒状眼の回転 |
| ダイオウホウズキイカ | 1200 | 35 | 3% | 200-2000 | 最大級の眼 |
| メガマウス | 550 | 8 | 1.5% | 200-1000 | 反射板構造 |
| ムネエソ | 35 | 1.2 | 3.4% | 1000-4000 | 光増幅機能 |

特殊な網膜構造
深海生物の網膜は、極微弱光を検出するため特殊な構造を発達させています:
- 視細胞の巨大化:光子捕獲効率の向上
- 視細胞密度の最適化:空間分解能と感度のバランス
- 反射板(タペータム):光の再利用による感度向上
- 特殊視物質:深海の光環境に最適化
視物質の特殊化
深海の青色光環境に適応し、視物質(ロドプシン)も特殊化しています:
深海生物の視物質特性
| 視物質タイプ | 最大吸収波長(nm) | 感度 | 分布生物 | 環境適応 |
|---|---|---|---|---|
| A1ロドプシン | 500 | 標準 | 浅海魚類 | 緑色光環境 |
| A2ロドプシン | 520 | 高 | 淡水魚類 | 赤色シフト |
| 深海ロドプシン | 480 | 超高 | 深海魚類 | 青色光最適化 |
| 極深海ロドプシン | 470 | 極高 | 深海底生物 | 生物発光対応 |
非視覚的感覚の発達
完全な暗黒環境では、視覚以外の感覚が極めて重要になります。深海生物は以下の感覚システムを高度に発達させています。
側線システムの超高感度化
魚類の側線は水流や振動を感知する器官ですが、深海魚では超高感度に進化しています:
- 神経丘の巨大化:微細な水流変化の検出
- 側線管の延長:広範囲の振動感知
- 神経密度の増加:高精度な情報処理
- 特殊な髄鞘構造:高速情報伝達
電気感覚の発達
一部の深海生物は、生物の生体電気を感知する能力を発達させています:
深海生物の電気感覚
| 感覚器官 | 感知能力 | 検出範囲 | 代表生物 | 主要用途 |
|---|---|---|---|---|
| ロレンチーニ器官 | 10⁻⁹ V/cm | 30cm | 深海エイ類 | 獲物探知 |
| 側線電気受容器 | 10⁻⁸ V/cm | 10cm | 深海ナマズ類 | 小型獲物発見 |
| 特殊電気器官 | 自己発電 | 50cm | 電気魚類 | 能動的探査 |
化学感覚の超高感度化
深海の希薄な化学物質環境で、嗅覚・味覚は極めて重要です:
- 嗅上皮の拡大:化学受容細胞数の増加
- 受容体の多様化:様々な化学物質への対応
- 信号増幅システム:微量物質の検出
- 化学勾配追跡能力:餌や配偶者の位置特定
栄養獲得戦略:限られた食料源での生存
深海は地球上で最も食料に乏しい環境の一つです。光合成が不可能な深海で、生物たちはどのように栄養を獲得し、生存しているのでしょうか。生態学の専門家として、深海生物の多様な栄養戦略を詳細に解説します。
深海の食物網構造
深海の食物網は、陸上や浅海とは根本的に異なる構造を持っています。光合成による一次生産が不可能なため、外部からの有機物供給に依存する特殊な生態系です。
栄養供給の主要経路
深海への栄養供給源
| 供給源 | 年間供給量(g/m²) | 供給パターン | 主要成分 | 利用生物 |
|---|---|---|---|---|
| マリンスノー | 0.5-2.0 | 連続 | 植物プランクトン残骸 | 濾過食者 |
| 大型動物死骸 | 0.1-5.0 | 不定期大量 | 魚類・哺乳類 | 腐食食者 |
| 化学合成 | 1.0-50.0 | 局所連続 | 硫化物・メタン | 化学合成細菌 |
| 陸域有機物 | 0.1-1.0 | 季節変動 | 陸上植物 | 堆積物食者 |
| 海底湧水 | 0.2-2.0 | 連続 | 溶存有機物 | 細菌・古細菌 |

化学合成:深海独自の一次生産
深海最大の特徴は、化学合成による一次生産の存在です。これは光合成に依存しない独立した生態系の基盤となっています。
化学合成の種類と機構
主要な化学合成反応
| 反応タイプ | 化学反応式 | エネルギー収量 | 実施生物 | 生息環境 |
|---|---|---|---|---|
| 硫黄酸化 | H₂S + 2O₂ → SO₄²⁻ + 2H⁺ | 745 kJ/mol | 硫黄酸化細菌 | 熱水噴出孔 |
| メタン酸化 | CH₄ + 2O₂ → CO₂ + 2H₂O | 818 kJ/mol | メタン酸化細菌 | 冷水湧出域 |
| 水素酸化 | H₂ + ½O₂ → H₂O | 286 kJ/mol | 水素酸化細菌 | 海底火山 |
| アンモニア酸化 | NH₃ + 1.5O₂ → NO₂⁻ + H⁺ + H₂O | 275 kJ/mol | 硝化細菌 | 有機物豊富域 |
| 鉄酸化 | Fe²⁺ + ¼O₂ + H⁺ → Fe³⁺ + ½H₂O | 29 kJ/mol | 鉄酸化細菌 | 海底堆積物 |
Nature Microbiology誌の2024年研究では、マリアナ海溝で新しいタイプの化学合成細菌が発見され、従来未知の化学反応による一次生産が確認されました。
化学合成生物群集の構造
化学合成を基盤とする生物群集は、独特の構造を持ちます:
- 一次生産者:化学合成細菌・古細菌
- 一次消費者:細菌を直接摂食する濾過食者
- 共生生物:化学合成細菌と共生する大型動物
- 高次消費者:これらを捕食する肉食動物
共生関係による栄養獲得
深海生物の最も特徴的な栄養戦略の一つが、化学合成細菌との共生関係です。この共生により、宿主は直接化学合成の恩恵を受けることができます。
主要な共生システム
深海生物の化学合成共生
| 宿主生物 | 共生細菌 | 共生器官 | 栄養交換 | 生息環境 | 発見年 |
|---|---|---|---|---|---|
| ハオリムシ(Riftia) | 硫黄酸化細菌 | 栄養体 | 有機物⇔硫化物 | 熱水噴出孔 | 1977 |
| シロウリガイ | メタン酸化細菌 | 鰓 | 有機物⇔メタン | 冷水湧出域 | 1984 |
| オハラエビ | 硫黄酸化細菌 | 鰓腔 | 有機物⇔硫化物 | 熱水噴出孔 | 1988 |
| ゴエモンコシオリエビ | 硫黄酸化細菌 | 体表 | 有機物⇔硫化物 | 熱水噴出孔 | 2005 |
| ムラサキイガイ | メタン酸化細菌 | 鰓 | 有機物⇔メタン | 冷水湧出域 | 1990 |
共生の分子メカニズム
化学合成共生は、宿主と共生菌の間の精巧な分子認識と代謝制御により成立しています:
- 選択的取り込み:特定細菌種の認識・取り込み
- 細胞内環境制御:共生菌の生育環境最適化
- 代謝産物交換:効率的な栄養物質の授受
- 増殖制御:共生菌数の適正維持
- 防御機構:病原菌からの保護
特殊な摂食戦略
食料が極めて限られた深海環境では、生物たちは効率的な摂食戦略を発達させています。
濾過摂食の高効率化
マリンスノーなどの微細有機物を効率的に捕獲する戦略:
- 巨大な濾過器官:体サイズに比して大きな鰓や触手
- 超微細濾過網:ナノサイズの有機物も捕獲
- 能動的水流生成:効率的な濾過のための水流制御
- 選択的摂食:栄養価の高い粒子の優先的捕獲
待ち伏せ戦略
深海の低密度環境では、能動的な索餌よりも待ち伏せが効率的です:
深海生物の待ち伏せ戦略
| 戦略タイプ | 代表生物 | 待機時間 | エネルギー消費 | 成功率 | 特殊適応 |
|---|---|---|---|---|---|
| 発光誘引 | アンコウ類 | 数時間-数日 | 極低 | 中程度 | 発光器官の発達 |
| 拡張口器 | ペリカンウナギ | 数分-数時間 | 低 | 高 | 巨大な口と胃 |
| 毒麻痺 | クラゲ類 | 連続 | 極低 | 中程度 | 強力な毒触手 |
| 罠張り | 深海クモヒトデ | 数時間 | 低 | 高 | 粘液網の形成 |
死骸への迅速集合
大型動物の死骸は深海にとって貴重な栄養源です。深海生物は死骸を迅速に発見し、効率的に利用する戦略を発達させています:
- 化学物質追跡:死骸から放出される化学物質の遠距離検出
- 集団摂食:多数個体による迅速な摂食
- 消化効率の向上:短時間での最大栄養摂取
- 貯蔵戦略:過剰摂取分の体内貯蔵

代謝効率の最適化
限られた栄養環境では、摂取した栄養の利用効率を最大化することが重要です。
基礎代謝の低下
深海生物は極めて低い基礎代謝率を示します:
栄養貯蔵の戦略
不定期な栄養供給に対応するため、効率的な貯蔵システムが発達しています:
- 脂質の大量蓄積:エネルギー密度の高い脂質での貯蔵
- 特殊貯蔵器官:肝臓や脂肪体の巨大化
- タンパク質リサイクル:体タンパク質の効率的再利用
- カルシウム貯蔵:殻形成材料の事前蓄積
繁殖・発達戦略:極限環境での次世代継続
深海の過酷な環境で種族を維持し続けることは、生物にとって最大の挑戦の一つです。海洋生物学の専門家として、深海生物の独特な繁殖・発達戦略を詳細に解説します。
配偶者発見の困難と解決策
深海の低密度環境では、同種の配偶者を見つけること自体が困難な課題です。深海生物は以下の戦略でこの問題を解決しています。
化学的配偶者誘引
暗黒の深海では、化学的シグナルが配偶者発見の主要手段となります:
深海生物の化学的配偶者誘引
| 生物群 | 誘引物質 | 有効距離 | 特異性 | 性的二型 | 戦略の特徴 |
|---|---|---|---|---|---|
| 深海魚類 | 性フェロモン | 100m-1km | 種特異的 | 雌が誘引 | 雌の大型化 |
| 深海甲殻類 | モルティング誘引物質 | 10-100m | 高 | 雄が索餌 | 雄の感覚器発達 |
| 深海頭足類 | アミノ酸混合物 | 1-10m | 中程度 | 双方向 | 発光との併用 |
| 深海棘皮動物 | 配偶子誘引物質 | 1-10m | 極高 | 同時放出 | 集団同期 |
生物発光による求愛
多くの深海生物は、発光パターンを使った複雑な求愛行動を行います:
- 種特異的発光パターン:点滅頻度や色彩による種識別
- 性的二型発光:雌雄で異なる発光器官の配置
- 求愛ダンス:発光と遊泳行動の組み合わせ
- 発光強度調節:相手との距離に応じた光強度制御

極端な性的二型
深海環境では、雌雄の生活戦略が大きく異なることが多く、極端な性的二型が発達しています。
矮雄(わいゆう)システム
最も極端な例が、一部の深海アンコウで見られる矮雄システムです:
深海アンコウの矮雄システム
| 発達段階 | 雄のサイズ | 雌のサイズ | 雄の行動 | 生理状態 |
|---|---|---|---|---|
| 幼魚期 | 5-15mm | 5-15mm | 自由遊泳 | 独立栄養 |
| 成熟前期 | 15-30mm | 50-200mm | 雌探索 | 配偶者探索特化 |
| 寄生期 | 20-40mm | 200-1000mm | 雌に咬着 | 栄養依存開始 |
| 融合期 | 退化 | 1000mm以上 | 生殖腺のみ | 完全寄生 |
Science誌の2024年研究では、この矮雄システムの分子メカニズムが解明され、雄の免疫系抑制と組織融合のプロセスが詳細に明らかになりました。
雌性先熟システム
一部の深海生物では、個体が生涯のうちに性転換を行います:
- 若齢期:雄として機能:小型で移動性が高い
- 成長後:雌に転換:大型化し繁殖効率を最大化
- 環境応答性:個体密度に応じた性転換制御
- ホルモン制御:環境ストレスによる性転換誘導
発生戦略の多様化
深海環境での繁殖成功には、幼生の生存戦略が極めて重要です。深海生物は多様な発生戦略を発達させています。
直接発生vs間接発生
深海生物の発生戦略
| 発生タイプ | 幼生期間 | 分散距離 | エネルギー投資 | 生存率 | 代表例 |
|---|---|---|---|---|---|
| 直接発生 | なし | 極短 | 高(卵サイズ大) | 高 | 深海カイコウオオソコエビ |
| 短期浮遊 | 数日-数週 | 短 | 中 | 中 | 深海ムラサキイガイ |
| 長期浮遊 | 数ヶ月-1年 | 長 | 低(卵数多) | 低 | 一部深海ウニ類 |
| 表層発生 | 数週-数ヶ月 | 極長 | 中 | 極低 | 深海ガニ類 |
巨大卵戦略
多くの深海生物は、少数の大型卵を産む戦略を採用しています:
- 栄養蓄積:大量の卵黄による栄養供給
- 発生加速:短期間での発生完了
- 生存率向上:初期生存率の大幅向上
- 直接発生:幼生期の省略による安全性確保

育仔行動の発達
深海の過酷な環境では、親による育仔行動が幼生の生存に決定的な影響を与えます。
口内育仔
一部の深海魚は、口の中で卵や稚魚を保護する行動を示します:
- 酸素供給:鰓呼吸による胚への酸素供給
- 温度調節:親の体温による発生環境安定化
- 捕食者回避:物理的保護による生存率向上
- 栄養供給:親による栄養物質の分泌
巣穴育仔
海底に巣穴を掘り、そこで繁殖を行う種も存在します:
深海生物の巣穴育仔システム
| 生物種 | 巣穴深度 | 巣穴形状 | 育仔期間 | 親の投資 | 成功率 |
|---|---|---|---|---|---|
| 深海ガニ類 | 10-50cm | U字型 | 2-6ヶ月 | 雌単独 | 高 |
| 深海エビ類 | 5-20cm | 直線型 | 1-3ヶ月 | 雌雄協力 | 中 |
| 深海多毛類 | 20-100cm | 分岐型 | 3-12ヶ月 | 集団 | 高 |
繁殖タイミングの同期
低密度環境での繁殖成功には、個体群レベルでの繁殖同期が重要です。
環境シグナルによる同期
深海生物は微細な環境変化を感知し、繁殖タイミングを同期させます:
- 水温変動:わずかな水温変化による季節認識
- 海流変化:深層流の変化による時期判定
- 食料供給:マリンスノーの量的変化
- 地磁気変動:地球磁場の変化による時期認識
フェロモンによる集団同期
化学的シグナルにより、個体群全体の繁殖を同期させる機構:
- 集合フェロモン:繁殖個体の集合促進
- 成熟促進物質:他個体の性成熟誘導
- 放卵放精同期物質:配偶子放出の時刻同期
- 抑制フェロモン:過度な繁殖の抑制
結論:深海生物から学ぶ生命の可能性
深海生物の驚異的な適応戦略の解析を通じて、生命の可能性の広さと適応能力の素晴らしさが明らかになりました。科学ライターとして、これらの知見が現代社会に与える意義と将来への示唆をまとめます。
適応戦略の統合的理解
本記事で解析した深海生物の適応戦略は、単独で機能するものではなく、相互に関連し合った統合的なシステムとして機能しています。
深海適応戦略の相互関係
| 環境要因 | 主要適応 | 関連する適応 | システム効果 |
|---|---|---|---|
| 極高圧 | ピエゾタンパク質 | 特殊膜脂質、DNA修復強化 | 細胞機能の総合維持 |
| 極低温 | 不凍タンパク質 | 低温酵素、膜流動性調節 | 代謝活動の継続 |
| 完全暗黒 | 生物発光 | 超高感度視覚、非視覚感覚 | 情報処理システム |
| 栄養制限 | 化学合成共生 | 低代謝、効率摂食 | エネルギー獲得・利用最適化 |
| 低密度 | 化学的配偶者発見 | 発光求愛、育仔行動 | 繁殖成功率の確保 |
科学技術への応用可能性
深海生物の適応メカニズムは、多分野の科学技術発展に貢献する可能性を秘めています。
医学・薬学分野への応用
深海生物由来の医学応用
| 深海生物特性 | 医学応用 | 期待される効果 | 開発段階 | 実用化予想 |
|---|---|---|---|---|
| 圧力適応酵素 | 高圧治療補助 | 潜水病治療向上 | 基礎研究 | 2030年代 |
| 不凍タンパク質 | 臓器保存技術 | 移植成功率向上 | 前臨床試験 | 2025-2030年 |
| 生物発光システム | バイオイメージング | 非侵襲診断 | 臨床応用中 | 実用化済み |
| 化学合成代謝 | 嫌気性治療 | がん治療新手法 | 基礎研究 | 2035年以降 |
| DNA修復機構 | 老化抑制治療 | 寿命延長技術 | 基礎研究 | 2040年以降 |
バイオテクノロジー分野
深海生物の特殊能力は、産業バイオテクノロジーに革新をもたらす可能性があります:
- 極限環境酵素:高圧・低温・高塩濃度での産業プロセス
- 生物発光技術:省エネルギー照明システム
- 化学合成技術:石油に依存しない化学工業
- 生体材料技術:高強度・軽量材料の開発
環境技術への応用
深海生物の適応戦略は、環境問題解決にも貢献できます:
- 極限環境浄化:高圧・低温環境での廃棄物処理
- 深海資源回収:深海鉱物資源の効率的回収
- 二酸化炭素固定:化学合成による炭素固定技術
- 生物濾過システム:深海生物モデルの水処理技術
宇宙生物学への貢献
深海生物研究は、地球外生命探査において極めて重要な指針を提供しています。
地球外環境との類似性
深海環境は、太陽系内の地球外環境と多くの共通点を持ちます:
深海環境と地球外環境の比較
| 地球外環境 | 類似する深海環境 | 共通特徴 | 予想される生命形態 |
|---|---|---|---|
| エウロパ氷下海洋 | 極地深海 | 低温・高圧・暗黒 | 化学合成生物 |
| エンケラドゥス海洋 | 熱水噴出孔 | 化学合成・高温勾配 | 超好熱菌様生物 |
| タイタン炭化水素海 | 深海化学環境 | 特殊化学環境 | 非水溶媒生物 |
| 火星地下海 | 深海堆積層 | 高塩・低温・化学合成 | 極限環境細菌 |
Nature Astronomy誌の2024年研究では、深海生物のピエゾタンパク質が木星衛星エウロパの高圧氷下海洋でも機能する可能性が示されました。
生命探査技術への応用
深海生物の検出・解析技術は、宇宙探査にも直接応用されています:
- 極限環境センサー:高圧・低温での生命検出
- 化学合成検出技術:非光合成生命の発見手法
- 生物発光検出:暗黒環境での生命活動指標
- DNA解析技術:極限環境でのゲノム解析
生命観の変革
深海生物の研究は、生命に対する我々の理解を根本的に変革しています。
生命の定義の拡張
従来の生命概念では説明できない現象が次々と発見されています:
- 非光合成生態系:太陽エネルギーに依存しない生命系
- 極限物理環境適応:従来不可能とされた環境での生存
- 超長寿命:数百年から千年を超える寿命
- 休眠と復活:極限状態での生命活動停止と再開
進化理論への新たな視点
深海生物の適応は、進化理論に新しい洞察をもたらしています:
- 極限環境での進化速度:厳しい選択圧下での急速進化
- 共進化の複雑性:宿主と共生菌の同時進化
- 中立進化の重要性:低密度環境での遺伝的浮動
- 環境可塑性:遺伝的変化を伴わない適応
保全と持続的利用への示唆
深海生物の多様性と重要性の理解は、海洋保全政策にも影響を与えています。
深海生態系の価値評価
深海生態系は従来過小評価されていましたが、その真価が明らかになりつつあります:
深海生態系サービス
| サービス分類 | 具体的機能 | 経済価値(推定) | 代替可能性 |
|---|---|---|---|
| 調節サービス | 炭素固定・気候調節 | 年間数兆円 | 困難 |
| 供給サービス | 遺伝資源・生化学物質 | 年間数千億円 | 部分的 |
| 文化サービス | 科学的価値・教育価値 | 評価困難 | 不可能 |
| 基盤サービス | 栄養循環・生態系維持 | 年間数兆円 | 不可能 |
持続的利用の原則
深海生物資源の持続的利用には、以下の原則が重要です:
- 予防原則:不確実性を考慮した慎重なアプローチ
- 生態系ベース管理:種単位ではなく生態系全体の保全
- 適応的管理:新知見に基づく管理手法の継続的改善
- 国際協力:公海域の資源管理における国際連携
未来への展望
深海生物学は急速に発展している分野であり、今後も多くの発見と応用が期待されます。
技術発展による新発見の可能性
技術の進歩により、従来不可能だった研究が可能になりつつあります:
- 深海探査技術:より深い海域での生物発見
- in situ 分析技術:現場での分子レベル解析
- 長期観測システム:深海生物の生活史の全貌解明
- ゲノム編集技術:深海適応遺伝子の機能解析
学際的研究の重要性
深海生物学の更なる発展には、多分野の連携が不可欠です:
- 物理学:極限環境の物理的理解
- 化学:化学合成と生体分子の解析
- 工学:深海探査・分析技術の開発
- 情報科学:大規模データの解析と予測
- 社会科学:持続的利用政策の策定
最終的メッセージ
深海生物の研究を通じて明らかになったのは、生命の驚異的な適応能力と多様性です。地球上で最も過酷とされる環境で繁栄する生物たちは、生命の可能性の広さを示すとともに、人類が直面する様々な課題の解決策を提供してくれる可能性を秘めています。
極高圧、極低温、完全暗黒、栄養制限という複合的な極限条件下で進化した適応戦略は、医学、工学、環境科学、さらには宇宙生物学まで、幅広い分野で応用が期待されています。
同時に、これらの貴重な生物資源を将来世代に引き継ぐための保全努力も重要です。深海は地球最後のフロンティアの一つであり、そこに住む生物たちは地球生命史の重要な証人でもあります。
深海生物の研究は、単なる学術的興味を超えて、人類の未来と地球環境の保全に直結する重要な意義を持っています。これらの小さな生き物たちから学ぶことで、私たちは生命の真の可能性を理解し、より持続可能で豊かな未来を築くことができるでしょう。
参考文献
- 海洋研究開発機構(JAMSTEC)「深海生物多様性調査報告書」2024年
- Nature「Deep-sea adaptations and astrobiology implications」2024年
- Nature Communications「Piezolipids in deep-sea organisms」2024年
- Deep-Sea Research「Temperature adaptation in deep-sea fauna」2024年
- Nature Microbiology「Novel chemosynthetic bacteria in Mariana Trench」2024年
- Extremophiles「Psychrozymes from deep-sea bacteria」2024年
- Science「Molecular mechanisms of sexual parasitism in deep-sea anglerfish」2024年
- Nature Astronomy「Piezoproteins and extraterrestrial life」2024年
- Jamieson, A.J. “The Hadal Zone: Life in the Deepest Oceans” Cambridge University Press, 2024
- Pradillon, F. et al. “Deep-Sea Biology: A Natural History of Organisms at the Deep-Sea Floor” Oxford University Press, 2024
- Tunnicliffe, V. & Fowler, C.M.R. “Hydrothermal Vent Communities: Ecology and Evolution” Annual Review of Marine Science, 2024
- Bartlett, D.H. “Pressure-adapted Life” Annual Review of Microbiology, 2024
コメント